Структура нуклеиновых кислот
Вернемся теперь к рассмотрению фибриллярных биополимеров. Нити нуклеиновых кислот привлекли к себе внимание исследователей более 50 лет назад. Однако для их успешного изучения техника выделения и химической характеристики должны были достигнуть определенного уровня, позволяющего получать хорошие волокна и ориентировать их.
Первые рентгеноструктурные исследования ДНК были выполнены Уилкинсом, Фрэнклин и их сотрудниками. Образцы волокон ДНК, полученные из различных источников, дают одинаковые рентгенограммы. Натриевая соль ДНК существует в виде двух модификаций А и В. А-форма кристаллическая, в ней содержится 75% воды, а В-форма менее упорядоченна, и содержание в ней воды выше. А- и В-формы обратимо переходят друг в друга.
Мы уже знаем, что для установления наличия спиральных структур по рентгенограммам волокон были разработаны общие правила. Уотсон и Крик использовали рентгенографические данные Уилкинсона и Фрэнклин, а также требование спаривания оснований. На базе этих данных они строили пространственные модели цепи молекулы. Модели собирали из отдельных металлических деталей, изображающих элементы структуры молекулы ДНК и напоминающих детский конструктор. С помощью этих моделей Уотсон и Крик показали, что В-форма натриевой соли ДНК представляет собой двойную спираль (рис. 10.22). Она состоит из двух правых спиралей, образуемых полинуклеотидными цепями (гл. 3, рис. 3.1).
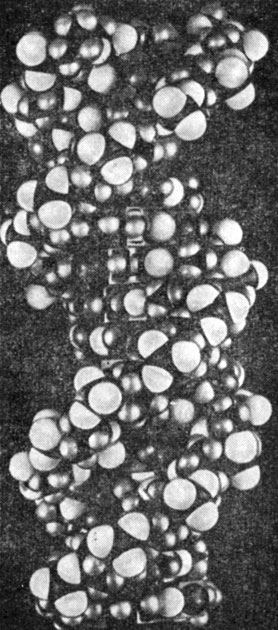
Рис. 10.22. Структура двойной спирали ДНК, предложенная Дж. Д. Уотсоном и Ф. Криком. Полинуклеотиды связаны друг с другом водородными связями, образующими поперечные мостики между пиримидиновыми и пуриновыми основаниями (рис. 3.1)
Фосфатные группы расположены на поверхности спирали, а основания находятся внутри. Плоскости расположенных друг над другом оснований перпендикулярны оси спирали. Расстояние по оси спирали между двумя парами относящихся к одной цепи оснований составляет 3,4 Å. На один виток укладывается 10 нуклеотидных остатков, и, следовательно, период спирали составляет 34 Å. Обе нити, образующие спираль, соединяют водородные связи между пуриновыми основаниями одной цепи и пиримидиновыми основаниями другой цепи - гуанин связывается с цитозином, а аденин - с тимином (гл. 3). Отношение парных оснований (аденина к тимину и гуанина к цитозину) почти всегда составляет единицу. Из этого следует, что последовательность расположения оснований в одной цепи автоматически определяет их последовательность в другой цепи. Это приводит к представлению о способе самовоспроизведения ДНК, который состоит в том, что цепи разделяются, а затем каждая из них служит матрицей, на которой образуется комплементарная цепь (рис. 10.23).
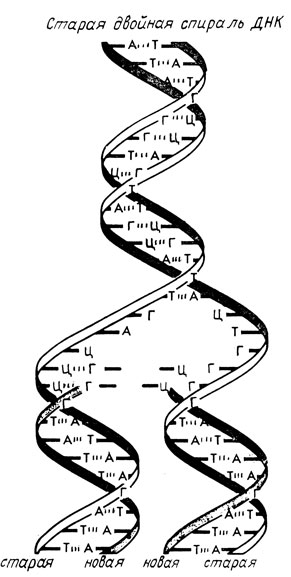
Рис. 10.23. Изображение механизма репликации ДНК. Новые нити ДНК образуются из трифосфатов, группируясь при помощи фермента ДНК-синтетазы в соответствии с родительской спиралью ДНК, которая служит матрицей
Показано, что А-форма также имеет структуру двойной спирали. Шаг спирали составляет 28 Å, а расстояние по вертикали между основаниями, относящимися к одной цепи, составляет 2,55 Å. Таким образом, на один виток укладывается 11 нуклеотидных остатков, а диаметр спирали составляет 18 Å. В этой форме ДНК плоскости оснований наклонены по отношению к оси спирали на угол 65°.
Механизм репликации ДНК, предложенный Уотсоном и Криком, был подтвержден открытием, сделанным Мезельсоном и Сталом. Бактерии, меченные 15N, помещали в среду, содержащую 14N. Через одно поколение из полностью меченных молекул образовались молекулы ДНК, меченные наполовину. Через два поколения бактерия содержала одинаковые количества наполовину меченной и немеченой ДНК. Мезельсон и Стал сумели разделить полностью меченную, наполовину меченную и немеченую ДНК по небольшому различию их плотностей, используя градиент плотности при ультрацентрифугировании. Эти результаты отчетливо показывают, что тяжелый азот в молекуле ДНК разделяется между двумя субъединицами и после репликации в каждой дочерней молекуле присутствует по одной субъединице из родительской молекулы ДНК (рис. 10.24).
![Рис. 10.24. Схематическое изображение поведения цепей ДНК, меченных 15N, при репликации в среде, содержащей строительный материал с 14N. [Meselson M., Stahl F. W., Proc. Natl. Acad. Sci., 44, 671 (1958)]](pic/000424.jpg)
Рис. 10.24. Схематическое изображение поведения цепей ДНК, меченных 15N, при репликации в среде, содержащей строительный материал с 14N. [Meselson M., Stahl F. W., Proc. Natl. Acad. Sci., 44, 671 (1958)]
Большая работа была проделана для установления структуры молекул РНК. Поскольку для этих молекул принцип спаривания оснований не выполняется строго, для большинства образцов РНК рибосом нельзя получить высококристаллические волокна. Рентгенографические исследования показали, что "транспортная РНК" имеет структуру, похожую на структуру А-формы ДНК. Двойную спираль образует одна цепь, сложенная вдвое. Большинство данных о структуре различных образцов РНК получено косвенным образом, при изучении спектров поглощения, светорассеяния, вязкости и ультрацентрифугирования растворов*.
* (Недавно была расшифрована полная трехмерная структура фенилаланиновой тРНК и найдено расположение всех нуклеотидов. Работа выполнена с монокристаллами тРНК и их тяжело атомными производными, т. е. тем же путем, каким Перутц, Кендрью и сотрудники расшифровали структуру глобулярных белков. - Прим. перев.)
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'