3.2. Биоэлектрические явления
Если проткнуть кончиком микроэлектрода (диаметром 0,5 мкм) мембрану крупной клетки, в частности (хотя и не обязательно) нервной или мышечной, можно зарегистрировать разность потенциалов относительно другого электрода, находящегося во внешней среде (рис. 58). Для уменьшения диффузионного потенциала микроэлектрод заполняют 3М раствором KCl. Трансмембранный потенциал (потенциал покоя) аксона и мышечной клетки составляет -60 и -90 мВ соответственно. При этом содержимое клетки оказывается электроотрицательным по отношению к внешней среде. Величина трансмембранного потенциала для разных клеток варьирует от -20 до -200 мВ. Клетки одного типа имеют постоянный характерный для них потенциал покоя. При регистрации в препаратах неповрежденных нервных и мышечных клеток трансмембранный потенциал сохраняется на постоянном уровне в течение многих часов.
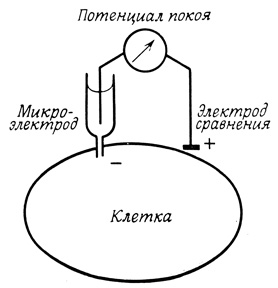
Рис. 58. Измерение потенциала покоя клетки
Разность потенциалов такого типа возникает на границе между двумя растворами электролитов, содержащих ионы с различной подвижностью в различных концентрациях. Нернст показал, что такой потенциал можно описать количественно следующим выражением:
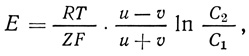
где R - универсальная газовая постоянная, Т - абсолютная температура, Z - валентность, F - число Фарадея, u - подвижность Na+, а v - подвижность Cl- для растворов NaCl с концентрациями С1 и С2.
В большинстве клеток функционирует система активного транспорта Na+/K+, которая поддерживает крутые градиенты концентраций этих ионов по обе стороны мембраны. Трансмембранный потенциал покоя представляет собой лишь следствие этого активного процесса. Фактически потенциал покоя зависит от различной подвижности ионов К+, Na+ и Cl-. Величина потенциала может быть выражена через концентрации этих ионов внутри (вн) и снаружи (сн) клетки и через коэффициент проницаемости ионов через мембрану (Р) в следующей форме уравнения Нернста:
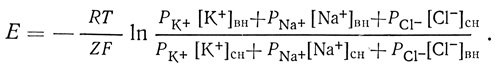
Мембрана аксона кальмара значительно более проницаема для К+, чем для Na+. По этой причине потенциал покоя для этого аксона зависит в основном от разности концентрации ионов К+:
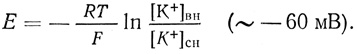
Это отчетливо доказывается тем, что при увеличении внешней концентрации ионов К+ потенциал покоя полностью исчезает, а при замене ионов Na+ на аналогично заряженные, но большие по размеру катионы холина практически не изменяется. Более высокая проницаемость мембран для ионов К+, чем для ионов Na+, связана, вероятно, с различными размерами гидратной оболочки этих ионов: диаметр гидратированного иона К+ меньше диаметра гидратированного иона Na+ (рис. 59).
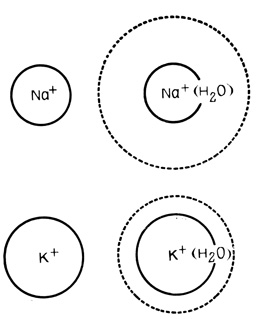
Рис. 59. Гидратные оболочки ионов К+ и Na+
Микроэлектроды могут быть использованы не только для измерения потенциала покоя, но и для пропускания тока с целью увеличения или уменьшения трансмембранного потенциала. Для изучения влияния таких изменений потенциала на электрические характеристики мембраны используют вторую пару электродов - регистрирующие электроды, - располагая ее на некотором расстоянии от первой пары (стимулирующие электроды, рис. 60).
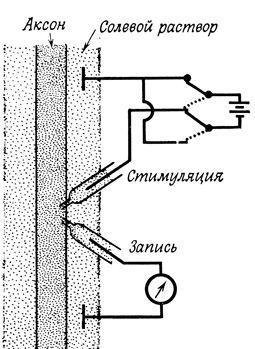
Рис. 60. Схема эксперимента по изучению влияния изменений трансмембранного потенциала на электрические характеристики мембраны
Короткий толчок тока соответствующей полярности, подаваемого через стимулирующий электрод, повышает отрицательный трансмембранный потенциал и вызывает кратковременную гиперполяризацию. Вызванный сдвиг быстро затухает, однако с помощью близко расположенных регистрирующих электродов его удается уловить. Характер затухания дает представление об электрических свойствах мембраны, например ее емкости, сопротивлении и т. д. Отрицательный потенциал можно увеличить в 2-4 раза по сравнению с потенциалом покоя, прежде чем произойдет электрическое повреждение мембраны.
Если ток, подаваемый через стимулирующий электрод, снижает исходный отрицательный потенциал (вызывает деполяризацию), то по достижении порога возникает кратковременный ток через мембрану, происходит "реверсия" трансмембранного потенциала - его знак меняется на обратный. Такая спонтанная реверсия, генерируемая самой мембраной, и представляет собой потенциал действия; в случае нервного волокна это электрическое возбуждение распространяется по мембране в форме незатухающего сигнала. Регистрирующие электроды улавливают мгновенную реверсию мембранного потенциала (в течение 0,5 мс) и последующее восстановление потенциала покоя (рис. 61).
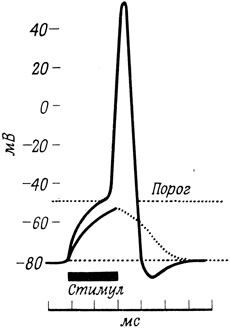
Рис. 61. Реверсия трансмембранного потенциала и возникновение потенциала действия
Потенциал действия длится всего несколько миллисекунд, после чего наступает так называемый рефрактерный период, который складывается из кратковременного интервала, в течение которого второй стимул - независимо от его силы - не способен вызвать потенциал действия, и периода постепенного, но быстрого восстановления. Повторные стимулы, наносимые по истечении рефрактерного периода, вызывают последовательные потенциалы действия (пиковые потенциалы, спайки); все они имеют одинаковую форму.
Описание потенциалов действия в терминах ионных потоков через мембрану стало возможным благодаря применению радиоактивных изотопов и метода фиксации напряжения. В последнем случае трансмембранный потенциал устанавливают на определенном уровне с помощью тока, который контролируется цепью с обратной связью. Если спонтанная реверсия потенциала не поддается контролю этим методом, то для фазы восстановления он вполне применим. Оказалось, что поток зарядов наружу происходит до тех пор, пока продолжается деполяризация.
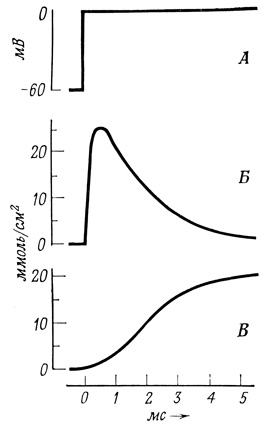
Рис. 62. Деполяризация и проводимость ионов. А - деполяризация; Б - проводимость Na+; В - проводимость К+
Спонтанная реверсия потенциала вызывается кратковременным поступлением ионов Na+ внутрь клетки, а восстановление потенциала покоя сопровождается более продолжительным выходом ионов К+ из клетки. На современном уровне наших знаний мы еще не можем описать на молекулярном уровне те каналы, по которым движутся эти ионы; можно лишь утверждать, что они специфичны и отличаются от обычных механизмов трансмембранного переноса ионов. Этот механизм ионной проводимости можно условно описать как своего рода "ворота". В таком случае можно представить себе, что потенциал действия возникает, когда в ответ на надпороговую деполяризацию на короткое время отворяются "ворота для натрия"; затем сразу же отворяются "ворота для калия", которые остаются открытыми до тех пор, пока не восстановится потенциал покоя (рис. 62 и 63). Величина тока относительно невелика (3⋅10-12-4⋅10-12 моль/см2), и его влияние на градиенты концентраций незначительно. Эти градиенты легко восстанавливаются благодаря работе Na+/K+-Hacoca.
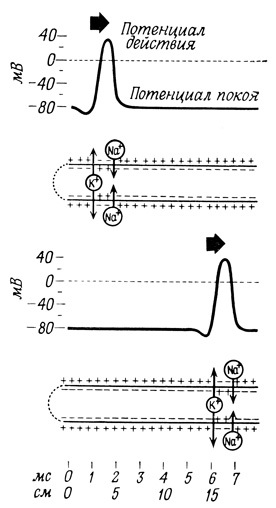
Рис. 63. Распространение потенциала действия по мембране и функционирование Na+/K+-нaсоса
Распространение импульса (потенциала действия) вдоль мембраны происходит оттого, что местные токи, возникающие в результате генерации потенциала действия, вызывают деполяризацию соседних точек на мембране. Скорость распространения импульса зависит от времени заряда емкости мембран. Время заряда определяется емкостью мембраны и величиной продольного сопротивления цитоплазмы аксона. Скорость распространения потенциала действия вдоль аксона зависит от его диаметра. У некоторых беспозвоночных быстрая реакция на стимул обеспечивается аксонами исключительно большого диаметра; например, гигантский аксон кальмара, иннервирующий его мантию, достигает в диаметре 1 мм.
В более сложно организованной нервной системе позвоночных распространению импульса по нервным волокнам способствует миелиновая оболочка аксона - изолирующий футляр, образованный клетками-сателлитами - шванновскими или глиальными. Цитоплазма шванновской клетки многократно обвивается вокруг волокна, формируя многослойную цилиндрическую "муфту". Мембрана аксона остается непокрытой только в области так называемых "перехватов Ранвье". Миелиновые сегменты обеспечивают более быструю и метаболически экономную передачу электрического сигнала по сравнению с "голым" аксоном (рис. 64).
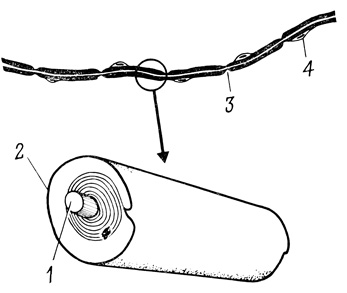
Рис. 64. Шванновская оболочка аксона. 1 - аксон; 2 - мембрана шванновской клетки; 3 - перехват Ранвье; 4 - ядро шванновской клетки
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'