6.2. Липидные компоненты

Рис. 111. Липидные молекулы содержат различные полярные 'головки' и длинные углеводородные остатки
Все мембранные липиды являются амфифильными молекулами. В состав липидных молекул входят, с одной стороны, длинные углеводородные остатки, отличающиеся высокой липофильностью и низким сродством к воде (гидрофобные), а с другой - более компактные гидрофильные полярные группы, получившие название полярных головок (рис. 111). Так, молекула глицерофосфолипида состоит из остатка глицерина, в котором две спиртовые группы (в положении 1 и 2) замещены ацильной или простой эфирной группами с длинными углеводородными цепями, образующими объемную липофильную область. В состав молекулы входит также полярная группа (полярная головка), включающая остаток фосфорной кислоты и образующая фосфоэфирную связь с третьей ОН-группой глицеринового остатка*. Структуры некоторых важных мембранных липидов приведены на рис. 112 и 113. Молекулы подобных соединений проявляют тенденцию к агрегации. При этом липофильные области молекулы, стремясь попасть в гидрофобную фазу, образуют непрерывные неполярные структуры, а полярные группы вместе с полярными группами других молекул формируют границу раздела фаз с водой. Геометрия возникающих липидных структур сильно зависит от природы входящих в состав компонентов.
* (Наряду с глицеролипидами в природе широко распространены липиды, у которых связь между полярной головкой и углеводородной цепью осуществляется с помощью этиленгликоля, пропандиола или другого двухатомного спирта (диольные липиды). В состав мембран диольные липиды с фосфорсодержащими или углеводными головка ми входят лишь в виде минорных компонентов (при более высоком содержании они вызывают лизис). Однако некоторые клетки способны накапливать значительные количества нейтральных диольных липидов, молекулы которых несут два углеводородных остатка. - Прим. ред.)
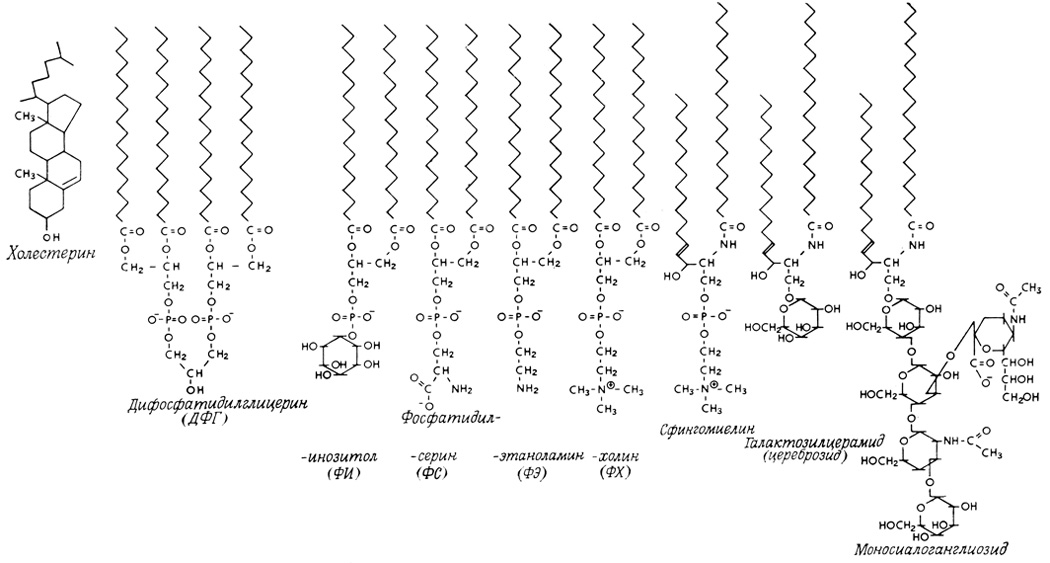
Рис. 112. Строение некоторых важных мембранных липидов
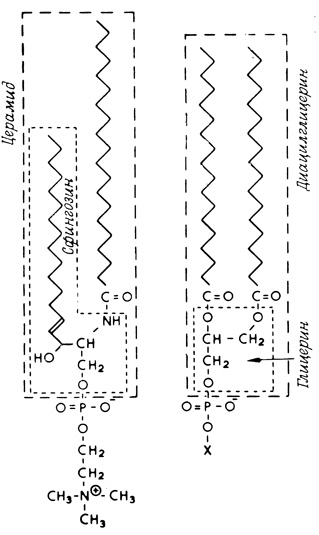
Рис. 113. Строение некоторых важных мембранных липидов
Некоторые липидные молекулы в мембранах можно заменить на другие или даже извлечь из мембран путем инкубации мембранных препаратов с водными суспензиями различных липидсодержащих агрегатов. Ряд липидов, в частности холестерин и слабополярные фосфолипиды, удается экстрагировать из мембраны органическими растворителями (скажем, эфиром или ацетоном), которые при низких температурах не вызывают существенных нарушений мембранной структуры. Однако большая часть полярных фосфолипидов экстрагируется из мембраны лишь с помощью системы растворителей, повреждающих молекулярную организацию мембраны (такова, например, смесь хлороформа и метанола). Возможность экстракции данного липида из мембраны зависит не только от локализации в мембранной структуре и сил, удерживающих липид в мембране, но и от сродства этого липида к соответствующему растворителю.
В мембранах животной клетки доминируют фосфолипиды. Однако некоторые мембраны, в частности поверхностные, содержат значительное количество нейтральных (менее полярных) липидов, главным образом холестерин и гликолипиды. Фосфолипиды представляют собой производные глицерина или сфингозина; помимо глицеринового или сфингозинового остатков, в молекулу такого липида входит полярная группа, содержащая остаток фосфорной кислоты, а также одна или две углеводородные цепи, связанные с остатком глицерина (сфингозина) сложноэфирной, амидной или эфирной связью. Различные фосфолипиды отличаются друг от друга как строением углеводородных цепей, так и природой полярных головок, однако фосфолипиды одного класса содержат, как правило, комплементарные ("спаренные") углеводородные цепи. Фосфолипиды настолько разнообразны, что даже в состав одной и той же мембраны входят обычно самые различные виды фосфолипидных молекул, которые, однако, удается сгруппировать в небольшое число классов. Гликолипиды животного происхождения - это главным образом производные сфингозина, которые столь же многообразны, как и фосфолипиды.
Липидный анализ многочисленных мембранных препаратов выявил поразительное сходство липидного состава однотипных мембран различных клеток. Значительная часть (от 35 до 75%) фосфолипидных молекул (ФХ, СМ и ЛФХ) клеточных мембран содержит цвиттерионную фосфорилхолиновую головку. В мембранах эритроцитов различных животных общее содержание ФХ и СМ остается сравнительно постоянным, а доля отдельных фосфолипидов варьирует в широких пределах. Мембраны животных клеток характеризуются относительно постоянным содержанием (от 15 до 25%) фосфолипидов, несущих большой отрицательный заряд (ФС, ФИ, ДФГ). Существенный вклад в суммарный отрицательный заряд мембраны при физиологических значениях pH вносит также ФЭ.
Липидный анализ препаратов плазматической мембраны различных животных клеток выявил высокое содержание в них холестерина; нередко оно равно общему содержанию фосфолипидов. Кроме того, эти мембраны сравнительно богаты гликолипидами. Мембраны комплекса Гольджи и вторичных лизосом также отличаются высоким содержанием холестерина, что, возможно, служит отражением особых взаимоотношений данных мембранных структур с плазматической мембраной при экзоцитозе и эндоцитозе (рис. 114). Мембраны указанных цитоплазматических органелл сходны с поверхностными мембранами и по содержанию СМ, которого нет в мембранах митохондрий и очень мало в ядерных мембранах и в эндоплазматической сети. ДФГ - фосфолипид, несущий большой отрицательный заряд, содержится в значительных количествах в мембранах митохондрий, но не входит в состав других мембран животной клетки*. Он является, однако, главным фосфолипидом бактериальных мембран, что послужило основанием для предположения о бактериальном происхождении митохондрий.
* (В некоторых опухолевых клетках наблюдается "выравнивание" липидного состава мембран различного типа. В таких клетках митохондрии могут содержать значительные количества СМ, а в мембранах эндоплазматической сети присутствует ДФГ. Это явление получило название "липидной дедифференцировки".- Прим. ред.)
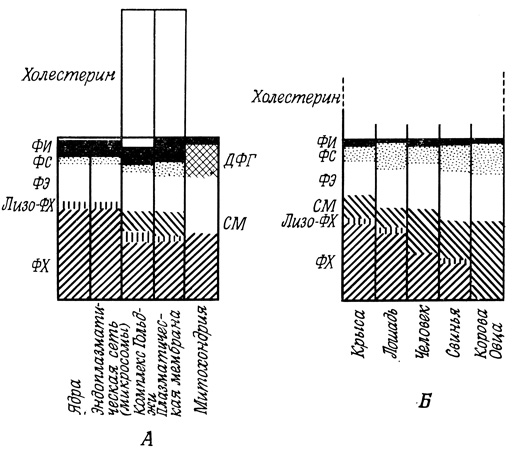
Рис. 114. Липидный анализ некоторых клеточных органелл и мембран в клетках печени (А) и состав мембран эритроцитов различных животных (Б). ФИ - фосфатидилинозит; ФС - фосфатидилсерин; ФЭ - фосфатидилэтаноламин; ФХ - фосфатидилхолин; ДФГ - дифосфатидилглицерин; СМ - сфингомиелин
Углеводородные остатки липидных молекул в мембранах животных клеток представляют собой неразветвленные цепи с четным числом атомов углерода, чаще всего от 14 до 24. Двойные связи в таких цепях (от 0 до 6 на цепь) имеют цис-конфигурацию. Мембраны некоторых цитоплазматических органелл отличаются более высоким содержанием ненасыщенных углеводородных цепей по сравнению с плазматической мембраной той же клетки. Это, по-видимому, отражает различия в фосфолипидном и гликолипидном составе между соответствующими мембранами. Получены данные о влиянии пищевого рациона на содержание и характер жирных кислот в составе мембранных липидов. Благодаря более высокому содержанию холестерина и одновременно низкому уровню ненасыщенных жирных кислот липидная фаза плазматической мембраны отличается большей упорядоченностью (жесткостью) по сравнению с липидами цитоплазматических органелл (рис. 115).
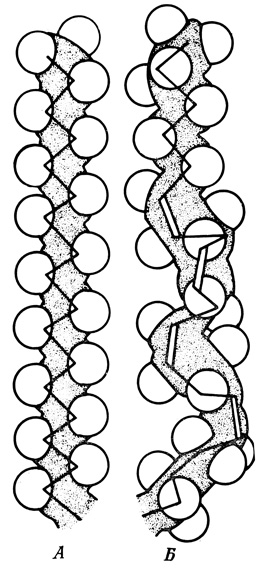
Рис. 115. Структура насыщенных (А) и ненасыщенных (Б) жирных кислот
В растительных клетках мембраны внутриклеточных органелл весьма сходны по липидному составу с соответствующими органеллами животных клеток. Хлоропласты же близки по липидному составу к сине-зеленым водорослям и, подобно последним, содержат моно- и дигалактозилдиглицериды, сульфогликозилдиглицериды и фосфатидилглицерин. Такое сходство может служить свидетельством в пользу гипотезы о происхождении хлоропластов из сине-зеленых водорослей. В состав мембраны хлоропласта входят также другие липиды (ФХ, ФЭ и ФИ), характерные для эукариотических клеток. Источником этих липидов служат, по-видимому, процессы липидного метаболизма, протекающие в других областях клетки (рис. 116).
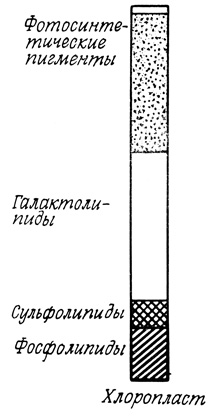
Рис. 116. Липиды, содержащиеся в мембранах растительных клеток
У дрожжей мембрана протопласта содержит около 40% липидов, которые представлены в основном нейтральными липидами и лишь 15-25% приходится на фосфолипиды. Главными компонентами последних являются ФЭ (33%) и ФХ (23%). Фосфолипиды с большим отрицательным зарядом (ФС, ФИ и ФА) составляют вместе почти 40%.
Липидный состав бактериальных мембран варьирует не только от штамма к штамму, но и в пределах одного штамма в зависимости от условий культивирования и фазы роста. Выделенные из бактериальных клеток мембранные фракции, содержащие плазматическую мембрану, мембраны лизосом, а иногда элементы клеточной стенки, содержат от 10 до 30% липидов. Более высокое содержание липидов было обнаружено в мембранах бактерий, лишенных клеточной стенки (L-формы), а также в мембранах Mycoplasma и Acholeplasma.
Фосфолипиды мембран грамположительных бактерий представлены в основном фосфатидилглицерином (ФГ) и ДФГ, а также аминоацильными производными ФГ. У грамотрицательных микроорганизмов в составе фосфолипидов мембран преобладает ФЭ. L-формы существенно отличаются по липидному составу от родительских клеток, причем этот состав у них в большей мере зависит от питательной среды. Например, холестерин среды включается в мембраны L-форм, даже если он полностью отсутствовал в родительской клетке. В обычных условиях холестерин является фактором роста для Mycoplasma, тогда как родственный последнему Acholeplasma, хотя и способен эффективно поглощать холестерин из среды, растет и в отсутствие этого соединения. Следует, правда, отметить, что при росте без холестерина клетки Acholeplasma имеют более хрупкие мембраны. Определенные штаммы Mycoplasma и Acholeplasma, а также некоторых бактерий могут включать жирные кислоты из культуральной среды в состав своих липидных молекул. Это позволяет произвольно изменять в эксперименте липидный состав исследуемых мембран. Было показано, что разветвленные углеводородные цепи, а также цепи с кольцевыми структурами вызывают дезорганизацию бактериальных липидов подобно тому, что происходит в липидах эукариотов под влиянием ненасыщенных двойных связей. В мембранах галофильных бактерий липиды представлены почти исключительно глицеролипидами, которые в качестве заместителей в положениях 2 и 3 (вместо обычных 1 и 2) содержат углеводородные цепи фитина, связанные с остатком глицерина простыми эфирными связями.
Часом симпатичные индивидуалки были готовы потешить тебя своими выдающимися возможностями в постели. Самые прелестные девушки со всего двора предлагают попользоваться к услугами интимного характера по разумным ценам. | Будьте одним из тех удачников, которые приобретут качественное прислуживание от самых очаровательных проституток. Всегда голые индивидуалки с вашего региона ежемесячно удовлетворяют молодых парней разными методами .
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'