Барьерные и коммуникационные свойства межклеточных контактов (Черненко Ю. П.)
Различные функции, выполняемые МК, сводятся к одной основной - системообразующей. Такие важные функции МК, как барьерная и коммуникативная, также целесообразно рассмотреть как аспекты системообразующей функции.
Клетки покровного эпителия и выстилающие различные полости организма (эпителиоциты, эпендимоциты, эндотелиоциты) соединены с помощью контактных структур в сплошные клеточные пласты. Области МК и обращенные в просвет плазмолеммы являются барьерами, препятствующими движению веществ через клеточные пласты либо обеспечивающими их селективный транспорт. Организмы животных, таким образом, разделены на ряд отсеков, или компартментов. Изоляция компартментов и коммуникации между ними необходимы для сохранения целостности организма и его жизнедеятельности. Поэтому исследование барьерных и транспортных свойств клеточных пластов является актуальной биологической и медицинской проблемой.
Существует два пути транспорта веществ через клеточные пласты: трансцеллюлярный - передвижение их через плазмолеммы и цитоплазму клеток (активный и пассивный транспорт, перенос в микровезикулах и везикулах) и парацеллюлярный - через области МК. Существует и смешанный путь - проникновение веществ в цитоплазму клеток и их последующий перенос в полость МК "в обход" барьерных контактных структур (рис. 13). Среди контактных структур барьерную функцию выполняют плотные соединения, а у беспозвоночных животных септированные. Барьерные и транспортные свойства плотных соединений, их морфофункциональная организация исследованы значительно полнее. Поэтому целесообразно остановиться на рассмотрении свойств этих соединений.
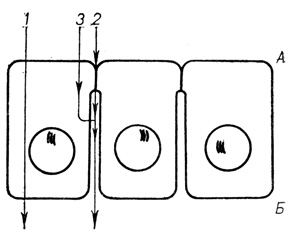
Рис. 13. Схематизированное изображение путей транспорта веществ через клеточный пласт: А - апикальные области, Б - базальные области клеток; 1 - трансцеллюлярный путь движения веществ, 2 - парацеллюлярный путь, 3 - смешанный путь движения веществ через цитоплазму клеток и межклеточные области
Как уже отмечалось, плотные соединения в большинстве случаев не являются идеальными межклеточными барьерами, препятствующими парацеллюлярному прохождению веществ, это своеобразные селекторы, избирательно пропускающие вещества в межклеточную область и через клеточный пласт. Сопротивление эпителиального пласта всегда меньше (часто во много раз), чем плазмолемм клеток. Единственной областью, шунтирующей величину трансэпителиального сопротивления, являются плотные соединения и парацеллюлярные пути. Если сопротивление плазмолеммы составляет 2900-4500 Ом/см2, то величина трансэпителиального сопротивления - 2000 Ом/см2. Это свидетельствует о том, что плотные соединения проницаемы для ионов Na+ и К+ (K. Kühn, E. Reale, 1975). Несмотря на общие свойства в структуре плотных соединений, существуют тканевые и органные особенности их ультраструктурной организации. Выделяют два основных типа плотных соединений; проницаемые, или "текущие" (leaky), и непроницаемые (tight) и соответственно этому проницаемые и плотные эпителии.
Эпителии с текущими плотными соединениями характеризуются низкими величинами трансэпителиального сопротивления - от 6 до 300 Ом/см2 (E. Fromter, 1972; P. Claude, D. A. Goodenough, 1973). К ним относят эпителий желчного пузыря, проксимальные извитые канальцы нефрона, солевые железы птиц. Плотные соединения в этих эпителиях представлены 1-4 линиями слияния наружных листков плазмолемм, линии слияния сплошные либо прерывистые (R. A. Ellis, 1977; В. Roesinger и др., 1978).
Плотные соединения между эпителиоцитами слизистой тонкой кишки, дистальных извитых канальцев нефрона относят к промежуточному типу, трансэпителиальное сопротивление при этом составляет 300-600 Ом/см2, т. е. во много раз ниже сопротивления апикальной плазмолеммы.
Эти соединения содержат до 8 линий слияния, формирующих сеть, которая сплошной поясовидной зоной окружает апикальные области клеток (beltlike tight junction). Такие соединения проницаемы только для мелких ионов и воды, более крупные молекулы (сахара, аминокислоты) транспортируются "в обход" плотных соединений. Вначале эти вещества переносятся через апикальную плазмолемму в цитоплазму клеток, а затем через латеральные плазмолеммы в межклеточные пространства (C. E. Stirling, 1972).
Применение микроэлектродного сканирования вдоль поверхности клеточного пласта позволило установить, что основными участками при прохождении электрического тока через эпителий являются межклеточные области. Около 96% электрического тока проходит по экстраклеточным путям, минуя клетки (E. Frömter, 1972). Ионы диффундируют по этим путям в гидратных оболочках (S. G. Schultz, 1977).
Непроницаемые плотные соединения обнаружены между эпителиоцитами слизистой оболочки желудка, мочевого пузыря, канальцев придатка яичка. Величина трансэпителиального сопротивления их может достигать 2000 Ом/см2. Непроницаемые плотные соединения имеют большую глубину, до нескольких микрометров, и состоят из большого числа линий слияния, например до 36 между сустентоцитами в семенниках крыс (D. S. Friend, N. В. Gilula, 1972; D. McGinley и др., 1972). Приведенные данные показывают, что ультраструктурная организация плотных соединений коррелирует с их свойствами: с увеличением числа линий слияния в плотных соединениях снижаются их транспортные свойства и нарастают барьерные. Эту закономерность можно проиллюстрировать на примере плотных соединений в нефроне. Транспортные процессы в нефроне наиболее выражены в проксимальных отделах, где через высокопроницаемые соединения и межклеточные пути из первичной мочи в кровь транспортируются неорганические ионы, микромолекулы и молекулы средних размеров. По мере нарастания вдистальном направлении нефрона осмотического давления мочи увеличивается и число линий слияния в плотных соединениях, возрастает их роль как межклеточных барьеров: в дистальных извитых канальцах и собирательных трубках эпителиоциты связаны непроницаемыми плотными соединениями (C. C. Tisher, W. E. Yarger, 1973; K. Kuhn, E. Reale, 1975).
Рассмотренные выше плотные соединения, за исключением проксимальных извитых канальцев нефрона, представляют собой сплошные поясовидные зоны. Наряду с этим существуют так называемые прерывистые, или макулярные, плотные соединения, в которых сетевидная область слияния наружных листков плазмолемм фрагментирована на разделенные промежутками участки (рис. 14). Такие плотные соединения представляют собой систему парацеллюлярных пор, или каналов, транспортные возможности этих соединений значительно шире. Так макулярные плотные соединения между мезотелиоцитами, эндотелиоцитами капилляров легких, поперечнополосатых мышц и сердца проницаемы для молекул пероксидазы хрена, молекулярная масса - 40 000 и микропероксидазы, молекулярная масса - 1900 (M. Simonescu, N. Simonescu, 1977; S. L. Wissig, M. C. Williams, 1978). Более крупные молекулы, например ферритин (диаметр молекулы 11 нм), через макулярные плотные соединения не проходят. Аналогичные результаты получены при исследовании проницаемости плотных соединений между эпендимоцитами, эндотелиоцитами капилляров радужной оболочки глаза, в корнеальном эндотелии (M. W. Brightman и др., 1970; N. Saari, 1975).
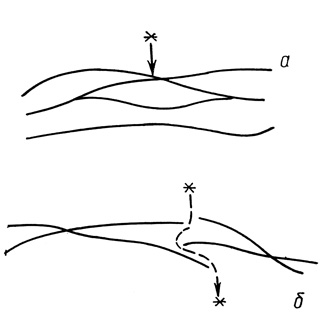
Рис. 14. Вид линий слияния наружных листков плазмолемм в сплошных, поясовидных, плотных соединениях (а), препятствующих прохождению веществ по парацеллюлярным путям и расположение линий слияния в фрагментированных плотных соединениях (б) у проницаемых для крупных молекул
Как видно из изложенного, ультраструктурная организация плотных соединений определяет барьерные и транспортные свойства клеточного пласта. В эндотелии сосудов плотные соединения между эндотелиоцитами являются важным компонентом гистогематических барьеров, от состояния которых зависит характер взаимоотношений между системами кровь - ткань. В капиллярах мозга, например, плотные соединения сплошного типа и проницаемы только для мелких ионов и воды, поэтому вещества, проходящие через стенки капилляров к нервным клеткам, "контролируются" цитоплазмой эндотелиоцитов. В органах, где плотные соединения между эндотелиоцитами кровеносных капилляров макулярного типа (легкие, сердце, поперечнополосатые мышцы), наблюдается парацеллюлярный переход веществ и перенос крупных белковых молекул через стенки капилляров.
Плотные соединения не являются статичными структурами, состояние их зависит от функционального состояния клеток и характера их жизнедеятельности, действия ряда экзогенных и эндогенных факторов. Так, в солевых железах птиц концентрация солевого секрета происходит за счет адсорбции из него воды, движущейся через плотные соединения между основными клетками. На высоте секреции глубина плотных соединений уменьшается до 10-40 нм (1-2 линии слияния), линии слияния становятся прерывистыми, т. е. происходит их фрагментация, перестройка сплошных плотных соединений в фокальные; в покое они снова приобретают вид сплошных, их глубина увеличивается до 30-90 нм (R. A. Ellis и др., 1977; C. V. Ridde, S. A. Ernst, 1977).
В кровеносных капиллярах плотные соединения на поверхности эндотелия занимают площадь всего 0,1%. Тем не менее модификация структуры плотных соединений может значительно изменять проницаемость сосудистой стенки. При действии гистамина или серотонина происходит макуляризация сплошных плотных соединений между эпителиоцитами. Это приводит к появлению парацеллюлярных гидрофильных каналов и возрастанию проницаемости капиллярной стенки (С. Flaxman и др., 1969; N. S. McNutt, R. S. Weinstein, 1973).
В стабилизации структуры плотных соединений значительную роль играют ионы Ca2+. Связывание внеклеточного Са2+ ЭГТА или ЭДТА приводит к дезорганизации и разрушению плотных соединений, что вызывает разъединение клеток. При этом резко падает сопротивление клеточного пласта. Через образовавшиеся межклеточные щели могут проходить крупные молекулы (P. Galli и др., 1976; M. Cereijido, 1978). Эти изменения обратимы, после добавления ионов Ca2+ ультраструктура и свойства плотных соединений нормализуются (J. Meldolesi и др., 1978). Восстановление ультраструктуры плотных соединений после декальцинирования может быть подавлено колхицином (J. Meldolesi и др., 1977). Под действием цитохалазина В, разрушающего цитоплазматические микрофиламенты, разъединяются клетки в участках плотных соединений (G. J. Kaye и др., 1974). Состояние цитоплазматических микрофиламентов и микротрубочек, таким образов, тесно связано со свойствами плотных соединений.
Дезорганизация и разрушение плотных соединений наблюдались при действии гиперосмотических растворов мочевины. Не-проницаемые в норме плотные соединения в эпителии мочевого пузыря амфибий под влиянием растворов мочевины резко повышают свою проницаемость (B. Wade-James и др., 1973). Обработка растворами мочевины кожи амфибий также приводит к резкому возрастанию проницаемости эпидермиса и падению трансэпителиального сопротивления (D. Erlij, A. Martinez-Palomo, 1972; S. Stender, 1973).
Под действием раствора мочевины проницаемость слизистой оболочки толстой кишки лишь незначительно повышается в отличие от тонкой кишки. Такую устойчивость плотных контактов в эпителии толстых кишок связывают с тем, что в естественных условиях в ней могут находиться токсичные для организма вещества (C. Storelli и др., 1977). Введение гиперосмотического раствора мочевины в кровоток головного мозга вызывает обратимое "открывание" гематоэнцефалического барьера (S. J. Rappoport, H. K. Thompson, 1973). Плотные соединения между эпителиоцитами желудка разрушаются под влиянием ацетилсалициловой кислоты (B. D. Frenning, 1972) и бактериальных токсинов. Так, сальмонеллезный эндотоксин приводит к макуляризации плотных соединений между энтероцитами и нарушению барьерных свойств кишечного эпителия (R. I. Walker, M. J. Porvaznik, 1978). Таким образом, микроорганизмы, обладая способностью деполимеризовать плотные соединения между клетками макроорганизмов, могут инъецировать через их клеточные барьеры свои метаболиты, занимая при этом новые клеточные территории.
Известны вещества, приводящие к разрастанию линий слияния в плотных соединениях и увеличению их глубины. Таким действием обладает, например, бициклический полипептид фаллоидин (R. Montesano и др., 1976). Следовательно, различные модификации структуры плотных соединений приводят к изменениям барьерных и транспортных свойств клеточных пластов и, очевидно, к перестройке коммуникационных процессов между различными компартментами организма. Нарушение структуры и свойств плотных соединений может быть важным патогенетическим звеном при развитии патологических процессов. При аллергическом энцефалите происходит дезорганизация плотных соединений между эндотелиоцитами капилляров мозга, через стенку таких капилляров проходят молекулы пероксидазы хрена (A. Hirano и др., 1970). Фрагментация плотных соединений может привести к возникновению аллергических заболеваний, в этих случаях молекулы антигенов или аутоантигенов могут проходить через клеточные пласты по парацеллюлярным путям в подлежащую соединительную ткань к иммунокомпетентным клеткам. Как аутоаллергический процесс можно объяснить мужскую стерильность (D. S. Friend, N. B. Gilula, 1972), различные пищевые и респираторные аллергии, дерматиты.
Одной из кардинальных функций МК является коммуникативная, благодаря которой осуществляется согласование процессов жизнедеятельности контактирующих клеток и их объединение в различные функциональные системы. Важность контактных взаимодействий клеток подчеркивалась еще О. Гертвигом (1900). Коммуникации между клетками обеспечиваются щелевыми соединениями посредством перехода различных молекул через гидрофильные каналы в этих соединениях. Однако коммуникативный процесс между клетками можно рассматривать шире. Изменения молекулярного состава и свойств контактирующих клеточных поверхностей, электростатических зарядов молекул и участков поверхностей, характер движения клеточной поверхности и ее структур и т. п. могут играть роль сигналов для соседних контактирующих клеток. С этой позиции контактные клеточные поверхности можно рассматривать как рецепторные поля, контролирующие состояние соседних клеток. В настоящее время коммуникативный процесс между клетками интерпретируется в рамках функциональной специализации щелевых соединений. Обнаружено структурное разнообразие щелевых соединений, что может указывать на различие их функциональных свойств (W. J. Larsen, 1977; С. Peracchia, 1977).
Через каналы щелевых соединений из клетки в клетку могут переходить неорганические ионы и различные микромолекулы, молекулы флуоресцирующих красителей, уридиновые и циклические нуклеотиды, нуклеиновые кислоты и низкомолекулярные белки (N. S. McNutt, 1974; C. Peracchia, 1977). О переносе неорганических ионов через щелевые соединения может свидетельствовать низкое электрическое сопротивление контактной области и большое значение коэффициента электрической связи между клетками, измеряемого как отношение величины мембранных потенциалов между двумя контактирующими клетками (VII VI ) при прохождении импульса электрического тока, приложенного к клетке I (W. R. Loewenstein, 1970). Коэффициент связи между контактирующими клетками в разных тканях и органах различен, в печени он составляет 0,6, между эпителиоцитами желудка - до 0,26, между тироцитами - 0,25-0,28, между клетками в культуре ткани - от 0,3 до 1,0 (W. R. Loewenstein, Y. Kanno, 1967; A. Jamakosmanovic, W. R. Loewenstein, 1968; Y. Kanno, Y. Matsui, 1968; D. F. Hulser, D. J. Webb, 1973). Контакты, проницаемые для ионов, проницаемы и для более крупных молекул. Ионные и метаболические связи обнаружены между клетками всех тканей. Таким образом, ткани представляют собой системы взаимодействующих клеток, своеобразные функциональные синцитии. Можно считать, что по аналогии с сосудистой сетью органа существует и тканевая контактная сеть, по которой метаболиты движутся непосредственно от клетки к клетке. В случае симметричной диффузии веществ через щелевые соединения ткани можно рассматривать как клеточные системы с усредненной внутренней средой. В действительности же клеточный состав ткани неоднороден и включает в себя камбиальные, дифференцирующиеся, активно работающие и резервные клетки, а также поврежденные, регенерирующие и элиминирующиеся клетки. Поэтому контактные взаимодействия тканевых клеток представляют собой чрезвычайно сложный процесс. Целесообразны исследования параметров пространственно-временной организации МК.
Ионная связь устанавливается при формировании щелевых соединений. Соприкосновение диспергированных клеток не приводит к немедленному ее установлению, но уже через 1-40 мин после контакта клетки начинают коммуницировать (W. R. Loewenstein, 1967, 1970; J. D. Sheridan, 1974; S. Но и др., 1977; S. Polak-Charcon и др., 1978).
Состояние коммуникативной функции зависит от клеточного метаболизма; охлаждение, действие разобщителей окислительного фосфорилирования, циклогексимида, актиномицина, повышение концентрации внутриклеточного Ca, повреждение клеток приводят к снижению коэффициента ионной связи между клетками или их полному функциональному разобщению (A. Politoff и др., 1967; R. J. Bushman, А. В. Taylor, 1971; S. Ito и др., 1977; С. Peracchia, 1978). При нарушении ионных связей между клетками нарушается межклеточный переход и других молекул, что свидетельствует о наличии общих транспортных путей для межклеточного переноса веществ. При локальном повреждении клетки в монослое уменьшается электрическая связь между ней и окружающими ее жизнеспособными клетками. Происходит своеобразная изоляция поврежденной клетки (P. O'Lague, H. Dalen, 1974). Такой механизм метаболического "отключения", по-видимому, препятствует попаданию метаболитов из поврежденных клеток в жизнеспособные. При различных нарушениях клеточного метаболизма ультраструктура щелевых соединений в ультратонких сечениях почти не изменяется, на криофрактограммах обнаружена незначительная дезорганизация внутримембранных частиц (С. Peracchia, 1978).
Межклеточные коммуникации и структуры, необходимые для их обеспечения, зависят от состояния генетического аппарата клеток и находятся под генетическим контролем. Так, если в культуре опухолевых клеток ионные связи между контактирующими клетками отсутствуют, то после слияния этих опухолевых клеток с нормальными образующиеся гибридные клетки устанавливают ионные связи, помимо ионов через контакты гибридных клеток могут проходить молекулы флуоресцеина, инъецированного в одну из контактирующих клеток (R. Azarnia, W. R. Loewenstein, 1973). Широко известен феномен метаболической кооперации между контактирующими клетками, при котором генетические дефекты в мутантных клетках корректируются при их контакте с нормальными клетками. Ультраструктурной основой для такого взаимодействия служат щелевые соединения, через которые из нормальных клеток в мутантные переносятся ферментативные продукты, нуклеотиды и их производные, перенос молекул РНК не обнаружен (H. Subak-Sharpe и др., 1969; R. P. Cox и др., 1972; N. B. Gilula и др., 1972; J. D. Potts, R. B. Burk, 1976; C. M. Corsaro, B. R. Migeon, 1977). Межклеточные коммуникации необходимы для реализации различных онтогенетических программ.
Благодаря контактным коммуникациям, каждая тканевая клетка находится под влиянием не только собственного генома, но и геномов соседних клеток. Действие геномов всех клеток популяции взаимно согласовано и имеет обобщенный, кооперативный характер (Г. Д. Туманишвили, 1968). Межклеточная коммуникация геномов между тканевыми, генетически однотипными клетками, представляет собой гомеостатический механизм для коррекции генетического дрейфа (А. Мак-Ларен, 1973).
Таким образом, коммуникации между клетками являются необходимым условием существования различных клеточных систем. Нарушения межклеточных коммуникаций могут лежать в основе развития патологических процессов. Исследования контактных взаимоотношений клеток в широком диапазоне клеточных функций помогут в решении задач практической и экспериментальной медицины.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'