14.3.4. Выход воды через устьица
Все три пути переноса воды завершаются испарением ее в воздушные пространства листа. Отсюда пары воды диффундируют через устьица по пути наименьшего сопротивления, т. е. от более высокого водного потенциала внутри листа к более низкому за его пределами. У двудольных растений устьица обычно располагаются в основном на нижней поверхности листа, на верхней же их меньше или нет вовсе. Регуляцию открывания и закрывания устьиц мы рассмотрим в разд. 14.3.9. Все листья окружены неподвижным слоем воздуха, толщина которого определяется размерами листа, особенностями его строения, например опущенностью, и скоростью ветра. Пары воды сначала диффундируют через этот слой и лишь затем уносятся воздушными потоками. Чем тоньше неподвижный слой, тем быстрее идет транспирация. Существует диффузионный градиент, направленный от неподвижного слоя назад к клеткам мезофилла. Теоретически каждое устьице должно иметь свой собственный диффузионный градиент - свою "диффузионную оболочку" (рис. 14.8). На самом же деле, если воздух неподвижен, диффузионные оболочки соседних устьиц перекрываются и образуют одну общую оболочку.
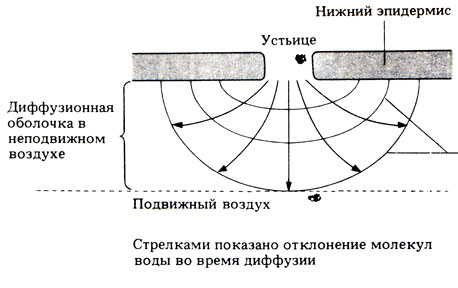
Рис. 14.8. Диффузия молекул воды из устьица. Обратите внимание на то, что у краев отверстия диффузионный градиент круче. Линиями показаны области одинаковой концентрации молекул воды (одинакового водного потенциала); чем круче градиент водного потенциала, тем ближе друг к другу расположены эти линии и тем быстрее происходит диффузия. Поэтому наибольшей будет скорость диффузии у краев устьичной щели. Этот 'краевой эффект' означает, что потеря влаги и газообмен будут более быстрыми при большом числе малых отверстий, чем при малом числе больших отверстий той же суммарной площади
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'